
Научная электронная библиотека
Монографии, изданные в издательстве Российской Академии Естествознания

Глава VII. СИСТЕМА ГИПОБИОМЕТАБОЛИЗМА У ПОЙКИЛОТЕРМНЫХ И ГЕТЕРОТЕРМНЫХ ОРГАНИЗМОВ
Использование льдоподобной формы воды, очевидно, потребовало соответствующих изменений в обмене веществ у организмов, во всяком случае, тех процессов, которые в своем завершении так или иначе связаны с молекулами воды. А в живом организме почти все системы имеют отношение к воде.
Как было уже показано, теплолюбивые растения, в т. ч. и ныне существующие, не приспособлены к использованию льдоподобной формы воды. При температурах ниже 4 °С, когда происходит физиологическая ангидрия они испытывают физиологическую сухость холодной почвы и криозасуху в их надземных органах, что является причиной гибели теплолюбов от холода. В отличие от них холодоустойчивые и засухоустойчивые растения, а также пойкилотермные и гетеротермные животные приспособились к льдоподобной воде и в определенные периоды жизни, в частности, во время гипобиоза используют ее для поддержания жизнедеятельности. Поэтому эти организмы могут без вреда перенести холод, то есть температуру с 4 °С до 0 °С. Организмы, способные перейти в состояние анабиоза могут частично использовать льдоподобную воду для синтетических процессов, направленных на образование ксеро- и криорезистентных структур и выдерживать уже значительные отрицательные температуры и отсутствие воды.
Каким же путем пошла эволюция организмов в использовании льдоподобной формы воды: путем повышения мобильности, пластичности уже имеющегося комплекса систем жизнедеятельности или путем создания качественно нового его аналога, имеющего сродство к льдоподобной воде?
В условиях теплого климата у древних растений и животных, очевидно, был теплолюбивый комплекс систем жизнедеятельности. Эту древнюю систему назовем архесистемой (от греч. arche – начало, начальный) жизнедеятельности, или для краткости архесистемой и обозначим буквами AСЖД. Архесистема является ликвофильной и сохраняет свою жизнеспособность до +4 °С – ниже уже инактивируется.
Нижней температурной границей жизнедеятельности истинных теплолюбов считается температура 4 °С. Значит архекомплекс систем жизнедеятельности у современных теплолюбивых растений и гомойотермных животных сохранился без изменения. Таким образом, интенсивность физиологических процессов (JT) у теплолюбивых организмов обеспечивается работой ликвофильной архесистемой жизнедеятельности:
JT = ACЖД.
Теперь рассмотрим систему жизнеобеспечения у пойкилотермных и гетеротермных организмов.
Как показали наши эксперименты [304, 305] холодоустойчивые растения, например, пшеница сорта «Якутянка-224» во время активной вегетации проявляют свойство теплолюбивости и при температурах от 4 °С до 0 °С на некоторое время находятся в состоянии криошока, выражающимся инактивацией физиологических процессов. Такое же явление наблюдается и у пойкилотермных животных организмов. Так живородящаяся ящерица (Lacerta vivipara), прекрасно зимующая в суровых условиях Якутии в летнее время не выдерживала даже температуру +5 °С и погибала [151]. Данное явление можно рассматривать с точки зрения биогенетического закона [144], как рекапитуляцию теплолюбивости с соответствующим типом водообмена у заведомо холодоустойчивых растений.
Таким образом, становится очевидным, что древняя предковая система жизнедеятельности – ACЖД у современных холодоустойчивых растений сохраняется полностью в неизменном виде и функционирует в теплое время, в период активной вегетации растений или активной жизнедеятельности других организмов. После холодового закаливания у пшеницы в области 4 °С можно уже наблюдать аномальную вспышку физиологических процессов. У других растений, например, у клубня картофеля происходит постепенное усиление интенсивности дыхания, которое начинается также с 4 °С [202]. При температуре ниже 4 °С происходит стратификация семян [199], закаливание растений и т. д. и т. п.
Как интерпретировать эти данные? Усиление дыхания клубней картофеля при температурах ниже 4 °С многие авторы объясняют повышением в их тканях содержания растворимых углеводов в результате интенсивного гидролиза крахмала [146, 181]. Есть еще одна гипотеза, объясняющая процесс яровизации семян [326]. Одной из характерных особенностей яровизации состоит в том, что она включает процессы, протекающие быстрее при низких температурах, чем при высоких [167]. Такая ситуация совершенно не характерна для химических процессов, поэтому, по мнению авторов, можно допустить, что изменения, происходящие во время яровизации в сущности представляют регулируемые ферментами реакции и проявляют все признаки таких реакций. Как тогда объяснить явный «отрицательный температурный коэффициент» яровизации у растений?
Для этой цели была выдвинута очень простая гипотеза, которая постулировала существование двух отдельных процессов, конкурирующих за общий субстрат. Каждый из процессов имеет положительные (хотя и различные) температурные коэффициенты:

В данной схеме реакции I и II конкурируют за общий промежуточный продукт – Б. Допустим, что реакция III несет более высокий температурный коэффициент, чем реакция I и II. Это означает, что высокая температура благоприятствует реакции III и больше (Б) будет участвовать в этой реакции (Р), поэтому будет образовываться мало В. Однако, когда температура заметно снизится, скорость РIII будет подавляться больше, чем РII (поскольку, по определению РIII более чувствительна к изменению температуры). Следовательно, положительная температура благоприятствует реакции II, в результате чего В будет накапливаться при низких, а не высоких температурах. Таким образом, процесс суммарного образования В имеет, по-видимому, отрицательный температурный коэффициент, хотя каждая из трех включенных реакций имеет в действительности положительный температурный коэффициент. Прямых данных, подтверждающих эту гипотезу нет, но, как указывают сами авторы, ценность ее заключается в том, что она показывает, «как процесс может протекать быстрее при более низкой температуре без нарушения естественных законов химических реакций». Гипотеза эта не объясняет аномалии физиолого-биохимических процессов у растений при температурах, близких к 4 °С, и другие особенности жизнедеятельности организмов в холоде.
Предлагаемая нами ниже гипотеза объясняет это явление c другой точки зрения, и опирается на имеющиеся прямые экспериментальные материалы.
Известно, что теплолюбивые растения нельзя сделать холодоустойчивыми путем холодового закаливания. Правда, при этом удается незначительно увеличить их резистентность к холоду, что, однако, происходит, по всей вероятности, за счет неспецифической реакции организма [292], а не в результате перестройки работы архекомплекса систем жизнедеятельности.
Из сказанного следует, что ACЖД обладает определенной консервативностью, не лабилен и не динамичен, поэтому не может нормально функционировать в среде льдоподобной воды, то есть при температурах ниже 4 °С, что с успехом осуществляется у холодоустойчивых растений. Скорее всего, в ходе эволюции у холодоустойчивых растений образовался новый комплекс систем жизнеобеспечения, дополнительно к уже существующему, то есть (АСЖД). Этот комплекс имеет сродство к льдоподобной воде и работоспособен в условиях холода и засухи. Данная гипотеза подкрепляется фактами, которых в последние годы становится все больше и больше. О них мы будем говорить чуть позже. Сейчас рассмотрим работу системы жизнеобеспечения у холодоустойчивых оорганизмов на примере растений. Она, очевидно, является более сложной, чем у теплолюбов.
Интенсивность физиологических процессов у холодоустойчивых растений (Jx) во время их активной вегетации, то есть в теплое время, обеспечивается, как и у теплолюбов, работой архесистемы жизне- деятельности:
Jx = ACЖД.
При охлаждении растения, когда температура падает ниже 4 °С, архесистема жизнедеятельности инактивируется, теплолюбивые организмы при этом погибают, а у холодоустойчивых включается новый комплекс систем жизнеобеспечения, его назовем Системой гипобиометаболизма – СГБМ, то есть системой работающей во время гипобиоза. У пойкилотермных и гетеротермных организмов в условиях холода (от 4 °С до 0 °С) и засухи интенсивность физиологических процессов, поддерживаемая СГБМ, очень низка и обеспечивает только их жизнеспособность (гипобиоз). Однако с понижением температуры ее активность не прекращается, а наоборот, усиливается, поэтому она не устойчива к морозу.
У организмов, способных к анабиозу, в том числе у морозоустойчивых растений, система жизнеобеспечения, можно полагать, существенно не отличается от такового у холодоустойчивых организмов. Ее можно рассматривать как модифицированную систему гипобиометаболизма – м-СГБМ. У этой системы синтетическая деятельность более выражена, чем у холодоустойчивых, и этот процесс довольно активно идет во время первой фазы закаливания, затем по мере ангидрии постепенно затухает, пока практически не прекратится. В связи с этим, у этих организмов во время холода синтезируются компоненты клетки, комплементарные к структуре жидкого и твердого льда и криопротекторы, выполняющие защитную функцию. При этом, как указано выше, через определенное время м-СГБМ полностью теряет активность и, таким образом, становится устойчивым к действию мороза.
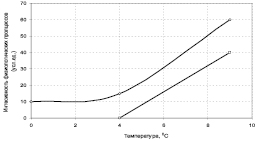
Рис. 7.1. Влияние закаливания на интенсивность физиологических процессов у растений: Верхняя линия – закаленные растения, работают две системы – JX = АСЖД + СГБМ; Нижняя линия – незакаленные растения, работает только одна система – JX = АСЖД
Системы гипобиометаболизма, в какой-то мере обладают терморезистентностью к повышенной температуре, так как в воде и при нормальной температуре сохраняется льдоподобная структура, которая поддерживает их нативность, что предполагает сохранение активности этих комплексов систем на некоторое время и после снятия холода, то есть при нормальных температурах. Следует заметить, что наличие льдоподобной воды при нормальной температуре обуславливает также возможность образования элементов СГБМ в этих условиях. Этим, например, можно объяснить некоторый закаливающий эффект температур, лежащих около 10 °С и ниже. Однако, полное включение СГБМ возможно только, начиная с 4 °С.
У закаленных холодом холодоустойчивых растений в условиях тепла будет работать 2 комплекса систем:
JX = АСЖД + СГБМ.
Интенсивность работы системы СГБМ по мере повышения температуры будет падать, чем выше температура, тем ниже их работоспособность. Благодаря совместной работе двух комплексов систем, интенсивность физиологических процессов у закаленных холодом растений будет значительно выше, чем у растений без такого закаливания (рис. 7.1). Эксперименты и практика показывают справедливость такого утверждения: умеренно закаленные холодом растения не только более жизнестойки, но и более продуктивны, чем контрольные растения [109, 140].
При неблагоприятных условиях существования, прежде всего, страдает энергетика растений [120]. Однако опыты, проведенные авторами, с использованием 2,4ДНФ показали, что усиление дыхания после холодового закаливания не сопровождается потерей энергетической эффективности дыхания.
Низкие ночные температуры до 10 °С оказывают благоприятное действие не только на рост стебля, но и интенсифицируют процессы цветения и плодоношения [161]. Предположение, что низкие ночные температуры приводят к повышению содержания сахаров в некоторых растениях в результате усиленного их перемещения, которое может дать эффект стимуляции, оказалось неверным. Экспериментальные данные свидетельствуют, что перемещение сахаров происходит наиболее интенсивно при 25 °С.
Было показано: добавка сахарозы извне не увеличивает дыхания кончиков корней гороха по сравнению с его исходной величиной [350]. Рост растения в растворе сахарозы также не увеличивался. Это, однако, не означает, что сахар не стимулирует интенсивность дыхания и ростовые процессы в меристематических и других тканях растений.
Проростки, закаленные холодом (+2 °С по 2,5 недели + –2 °С на 2 суток), морозостойкой пшеницы сорта «Одесская-16» имели почти в 1,5 раза более высокий уровень дыхания при 3 °С по сравнению с уровнем дыхания закаленных проростков маломорозостойкого сорта пшеницы «Ровенская 14». Незакаленные проростки обоих сортов существенно не отличались по интенсивности дыхания в исследованном диапазоне температур. В области высоких температур (35–45 °С) у сорта «Одесская 16» интенсивность дыхания проростков контрольного варианта выше, чем у варианта с закаленными растениями на 147 % [155]. Таким образом, эффект стимуляции холодового закаливания выше у более холодоустойчивых, чем у менее резистентных к холоду растений. Это вполне объяснимо так как, повышение синтетической активности у более холодоустойчивых растений происходит, благодаря одновременной работе двух систем – АСЖД и СГБМ.
Стимулирующее действие холодового закаливания семян и проростков в настоящее время общепризнанно и находит применение на практике.
Вероятно, первым изученным биохимическим процессом, ускоряющимся при пониженных температурах, был гидролиз крахмала в клубнях картофеля, в результате которой, в тканях накапливаются растворимые сахара [374]. В образовании сахаров участвует фермент – сахарасинтетаза. Например, при температуре +4 °С в проростках пшеницы на протяжении 14 дней наблюдалось увеличение (5–6-кратная индукция) количества сахарасинтетазы [353].
В последнее время найдено большое количество биохимических соединений, синтезирующихся при пониженных температурах и проявляющие высокую активность в холодных условиях. Так при пониженных температурах синтезируются стресс-белки [63, 64, 157, 343], меняется изоферментный состав белков [9, 193], накапливается АБК [290, 364], повышается ненасыщенность жирных кислот [43, 344] и другие вещества.
По данным В.К. Войникова и М.В. Корытова [64] закаливание проростков к холоду приводит к синтезу новых полипептидов. В течение первых суток гипотермии синтезируется 7 стрессовых белков. Через 2–3 суток гипотермии число стрессовых белков увеличивается почти в 3 раза. При дальнейшем закаливании проростков к холоду число стрессовых полипептидов сохраняется и к 7–11 суткам гипотермии оставшиеся в спектре белки формируют пул белков адаптации, состоящегося только из стресс-белков.
Серьезным аргументом в пользу наличия у холодоустойчивых растений качественно нового комплекса систем жизнеобеспечения, способного работать при пониженных температурах, является обнаруженная исследователями смена ферментных систем, а также переход дыхания при неблагоприятных условиях на другой путь, например, на пентозофосфатный [244, 254] или анаэробный тип дыхания у озимых и древесных растений [201].
Б.А. Рубин и М.Е. Дадыгина [245] пишут, что «… у зеленых растений каждый этап дыхания осуществляется при участии не какого-либо одного, а двух или большего числа ферментов, выполняющих зачастую одну и ту же или, во всяком случае, близкую функцию. Это служит убедительным свидетельством того, что окислительная система у этих организмов построена по принципу множественности (мультипринцип). С эволюционной точки зрения целесообразность одновременного присутствия в клетке нескольких ферментов, катализирующих одну и ту же химическую реакцию, может состоять лишь в том, что отдельные представители этого комплекса различаются по характеру зависимости их действия от условий среды. Это положение, непосредственно вытекающее из закона единства организма и среды, получило в настоящее время существенное экспериментальное обоснование» (с. 259–260).